|
||||
|
|
Глава перваяЭволюция Возникновение сложности Великая идея: эволюция идет путем естественного отбора Жизнь столь совершенна, что, как долгое время считали, ее было необходимо сотворить особо. Ибо как может нечто столь удивительное и столь уникальное самопроизвольно возникнуть из безжизненной слизи? В самом деле, что является той главной частью вещей, которая наделяет их жизнью? Ответы на эти и другие вопросы первостепенной важности появились двумя волнами. Первой была волна эмпирических объяснений, когда наблюдатели, по большей части натуралисты и геологи девятнадцатого века, пристально изучали внешние формы природы и получали далеко идущие выводы. Затем пришла вторая волна, в двадцатом веке, когда кроты с глазами ученых рыли ходы под поверхностью явлений и открывали молекулярную основу паутины жизни. Первый из этих подходов является предметом настоящей главы; второй, чрезвычайно обогативший наше понимание того, что значит быть живым, есть предмет следующей. Древнегреческие философы, как обычно, имели свои собственные взгляды на природу живых существ. Как и большинство их мнений, высказанных из самых лучших побуждений, эти взгляды были нелепо, но обаятельно превратны. Например, самопровозглашенный бог Эмпедокл (490-430 до н.э.), незадолго до принятия им не слишком мудрого решения продемонстрировать свою божественность путем низвержения себя в кратер вулкана Этна, предположил, что животные построены из универсального набора частей, которые, будучи собраны в различные комбинации, дают слона, комара, рогатую жабу и человека. Мир заселяется этими знакомыми комбинациями охотнее, чем летающими свиньями и ослами с рыбьим хвостом потому, что лишь некоторые комбинации жизнеспособны. Природа, вероятно, экспериментировала с другими комбинациями, в ожидании Острова доктора Моро, но после недолгого хромания, трепыхания и переваливания с боку на бок эти экспериментальные творения умирали. Почти на два тысячелетия позже эхом откликнулся похожий взгляд, но уже на молекулярном уровне. Его высказал граф Жорж-Луи Леклерк де Бюффон (1707-88), считавший, что организмы самопроизвольно возникли из агрегаций, которые мы сегодня назвали бы органическими молекулами, и что число возможных видов есть число жизнеспособных комбинаций этих молекул. Бюффон полагал, что уж он-то знает: свой великий труд Всеобщая и частная естественная история (Histoire naturelle, générale, et particulière), начатый в 1749 г., он планировал довести до пятидесяти томов, но успел подготовить тридцать шесть. Девять были посвящены птицам, пять минералам и восемь (опубликованных посмертно) китообразным, рептилиям и рыбам. Но откуда все эти творения, все живые существа действительно появились, их внушающее благоговение количество, два миллиона зарегистрированных видов и, возможно, десять и более миллионов, которые еще предстоит обнаружить? Аристотель, всегда блистательно обильный интеллектуально и, как всегда, блистательно ошибавшийся, предположил, что животные упали со звезд или произошли самопроизвольно уже в завершенном виде. Индейцы яхуна из бассейна Амазонки приняли неоаристотелевский взгляд и считали, что маниока выросла из пепла убитого и кремированного Миломаки. Индейцы кауилла из Калифорнии также верили, что виноградная лоза выросла из его кремированного живота, арбузы из зрачков его глаз, а злаки из его зубов. Менее общепринято мнение, что пшеница произошла из яиц его вшей, а бобы из его спермы. Другие религии предложили с виду простые мнения, согласно которым все существа, большие и малые, были сотворены Богом, и это все. Однако даже некоторые отцы церкви находили затруднительным примириться со всеми утверждениями Библии. Например, эрудит Григорий Назианин (330-89, Назиан был где-то в Каппадокии, в Малой Азии), считал, что Бог должен был создать некоторые из своих творений после потопа, поскольку небольшой ковчег Ноя был слишком мал для того, чтобы приютить представителей всех видов.[1] Архидиакон Карлайла Уильям Пэйли считал бесспорным, что он установил происхождение творений в книге с игривым названием Естественная теология или свидетельства существования и атрибуты божества, собранные из природных явлений, опубликованной в 1802 г., где в качестве превосходного аргумента он использует аналогию с путешественником, который столкнулся с ручными часами, созерцает их сложный замысел и не сомневается в том, что за этим замыслом стоит часовщик. Таким образом, каждый, кто сталкивается со сложностью природы должен неизбежно заключить, что к ее замыслу и конструкции приложил руку Бог. Однако Анаксимандр из Милета (610-545 до н.э.), делая вклад в западную философию, когда она была еще едва пробившимся зеленым побегом, действительно имел проблеск чего-то, похожего на истину. В совместной с Фалесом и Анаксименом философской программе он сделал чисто умозрительное заключение относительно всех живых существ и жизни в целом, гласящее, что виды животных могут превращаться один в другой. В науке часто бывает, что первым шагом к действительному пониманию, приходящему на смену фантастическим спекуляциям, оказывается собирание данных. В данном случае для этого надо идентифицировать и классифицировать все типы организмов, составляющих биосферу, или, по крайней мере, столько, сколько позволят терпение, настойчивость и провидение. Наиболее полезными бывают названия, выражающие родство, похожие на обычай давать членам одной семьи одну фамилию. К середине восемнадцатого века, когда установилась международная морская торговля, даже домоседы стали осведомленными об изобилии организмов и странных существ, населяющих мир, и поняли, что простых названий, таких как корова и собака, уже недостаточно. Это было подобно тому, как обитатели Лапландии вдруг обнаружили бы, что в Уганде недостаточно знания их родного языка. Первую общепризнанную систему наименований разработал шведский ботаник Карл фон Линней (Carl von Linne, 1707-78), в латинском написании Линнеус (Linnaeus). Линней изложил свою систему наименований в труде Systema naturae, опубликованном в 1735 г., а для извлечения на свет систематической классификации растений обычно обращаются к его книге Spesies plantarum, 1753. В этих работах Линней ввел иерархию групп (рис. 1.1) с царствами около вершины и пирамидой, расширяющейся ко все более частным разделам по мере нисхождения через типы, классы, отряды, семейства, роды и виды. Эта схема с тех пор пополнялась посредством включения различных промежуточных слоев, таких как подсемейство и надсемейство. Так, нас, людей, следует классифицировать (иронически — возразит кто-то) как вид Homo sapiens, рода Homo, в семействе Гоминидов, в надсемействе Гоминоидов инфраотряда Узконосых подотряда Антропоидов отряда Приматов подкласса Плацентарных в классе Млекопитающих, в надклассе Четвероногих из типа Хордовых в царстве Животных домена Ядерных в империи организмов. 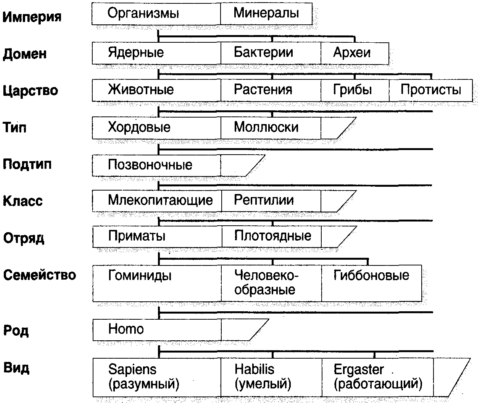
Недостатком системы Линнея является то, что она основана на внешне узнаваемых подобиях скорее, чем на более приемлемом в научном отношении установлении лежащего в их основе родства. Более того, точному определению классов, типов и т. д. трудно приписать, а в действительности и вовсе не удается какой-либо особенно глубокий фундаментальный смысл. Современным методом таксономии является кладистика (кладос, по-гречески молодой побег), тщательно исследующая происхождение организмов от общего предка и идентифицирующая различные ветви, или клады, дерева жизни (рис. 1.2). Кладйстика была введена немецким таксономистом Вилли Хеннигом (1913-1976) и доработана в его Филогенетической систематике (1966). Согласно Хеннигу, классификация должна отражать генеалогическое родство, и организмы должны группироваться строго на основе их происхождения от общего предка. Не в пример легкомысленным теоретическим физикам, приспособившим к своим схемам повседневные слова, такие как «спин» (в переводе с английского «вращение») и «аромат», Хенниг перегрузил таксономию греческими терминами, и кладисты имеют дело с симплециоморфами (характеристиками, разделяемыми более чем одной тварью), синапоморфами (разделяемыми производными характеристиками) и т.д. К счастью, у нас нет необходимости использовать этот перегруженный язык, поскольку мы главным образом будем пользоваться системой Линнея. Однако кладистика весьма влиятельна, логична и полезна, так как она основана на генеалогии организмов, которая, как можно доказать, есть единственный рациональный фундамент классификации. 
Роковое вымирание динозавров в конце Мелового периода — только наиболее известное из по крайней мере пяти главных событий. Среди когда-либо произошедших катастрофических событий находится то, которое привело к концу Пермский период (Пермь, в восточной части европейской России), с исчезновением более 95 процентов видов морских животных. Ордовикский период был резко прерван 440 миллионов лет назад, Девонский — 350 миллионов лет назад, Пермский период — 250 миллионов лет назад, Триас — 205 миллионов лет назад и Меловой — 65 миллионов лет назад. Причины большинства этих вымираний все еще по большей части неизвестны, но недостатка в идеях нет, включая столкновения с астероидами и большие падения уровня моря, сопровождавшиеся глобальным похолоданием. Сопутствующие вымирания являются травматическими, но жизнь не унывает, и многообразие видов возвращается очень быстро: через 5-10 миллионов лет многообразие достигает уровня, имевшегося до вымирания, часто даже превосходя его. События вымирания сметают конкурентов, освобождают ниши, созревшие для колонизации, и поздравляют всех (кроме вымерших) с появлением благоприятных возможностей. Однако, хотя вымирания играют важную роль, не следует ее преувеличивать. Типичный для животных вид продолжает существовать около 2 миллионов лет, а вспышки вымирания случаются обычно каждые 20-30 миллионов лет, так что большая часть видов не подвергается вымиранию от катастроф. Злосчастная судьба динозавров таилась в том, что они оказались слишком живучими: они жили достаточно долго и дождались. В настоящее время мы, по-видимому, находимся в сердцевине массового вымирания нового типа, когда активность человека делает биосферу неблагоприятной для многих видов флоры и фауны, с которыми он делит ее, а возможно, и для него самого. Самоиндуцируемое вымирание этого типа может быть неотвратимым спутником «прогресса», так как с ультрапессимистической неомальтузианской точки зрения можно считать, что способность к самоуничтожению неизбежно обгоняет развитие интеллекта. Наиболее мрачный взгляд заключается в том, что общества еще могут выживать, если индивид способен убить одним ударом всего несколько тысяч человек (как это было до сих пор в человеческой истории), но ни одно общество выжить не может, когда технология развита до такой степени, что одна персона обладает мощью, достаточной для убийства десятков миллионов. Человеческое общество, возможно, как раз подошло к этой точке. Если это является общим правилом для населения всех планет, то у нас очень мало надежды, так художественно внушаемой оптимистичной научной фантастикой, надежды на исполнение космических чаяний человечества. Зато наше вымирание, по крайней мере, создаст благоприятные возможности для тараканов. Остается несколько центральных вопросов, касающихся всей этой богатой взаимосвязи между географией и генами, этой танцплощадки естественного отбора. Один из них — о природе объекта, на который воздействует естественный отбор. Воздействует ли он на ген, на индивида или на вид? Мы можем исключить вид как единицу отбора. Организмы ничего не делают ради своего вида. Так же как естественный отбор не видит будущего, он не видит и сообществ. Индивид конкурирует с другим индивидом и стремится к своему собственному успеху, не заботясь о пользе совокупности организмов, составляющей вид. Репродуктивная энергия индивида питает эгоистическое поведение и не имеет представления об альтруизме, бессознательном поведении, приводящем к самопожертвованию ради других. Нельзя отрицать, что многие виды поведения выглядят альтруистическими, и лишь когда мы более тщательно исследуем их, обнаруживается, что это волки в овечьих шкурах, и альтруизм на деле оборачивается красными каплями эгоизма на зубах и клыках. При взаимном альтруизме, представляющем собой управляющий идеальным человеческим обществом вариант социального контракта, организм получает прощение за свой эгоизм в обмен на сотрудничество с другими организмами в значительной мере для того, чтобы в тяжелые времена помогающий мог бы сам получить помощь. На более глубоком уровне следует понимать, что представители вида имеют общие гены, и помогая очевидным конкурентам по размножению, организм скрыто помогает распространению своих собственных генов. Этот тип альтруизма назван родственным отбором. Таким способом биолог-теоретик Дж.Б.С. Холдейн (1892-1964) выразил точку зрения, согласно которой он был бы рад утонуть, если бы это спасло двух его родных или десять двоюродных братьев. Каждый из его родных братьев разделял бы с ним половину его генов; его двоюродные братья разделяли бы одну восьмую (для компенсации хватило бы спасения восьми двоюродных братьев, спасение десяти дает уже генное преимущество). Контроль наших генов над нашим поведением предполагает, что мы можем смотреть глубже, чем уровень вида, глубже чем уровень индивида, прямо в глубь генов. Проблема этого взгляда состоит в том, что он редко находится во взаимно однозначном соответствии с поведением. Существует не только заговор сложности биосферы, но также и воплощение генотипа (генетической конструкции организма) в фенотипе (физических характеристиках организма). Некоторые организмы отказывают себе в радостях воспроизводства, но все же делают свой вклад в будущее, помогая вместо себя размножаться своим близким родственникам. Гены их матки (например, пчелиной) так близки к их собственным, что, помогая ей размножаться вместо себя, они достигают распространения и своих генов: ведь она может рассеивать копии их генов, освобождая их от необходимости самим беспокоиться об этом. Другой проблемой является отслеживание последствий конкуренции на одном уровне (например, индивидуальном) для другого уровня (видового). Может случиться, что выгода индивида вредоносна для группы. Поскольку индивид не обладает эволюционным предвидением, он может пренебрегать последствиями своего поведения для группы. Когда пищи недостаточно, некоторые индивиды продолжают размножаться и передают свои гены следующим поколениям; они не воздерживаются ради своего вида. В результате вид будет эволюционировать в направлении, задаваемом генным потоком эгоистичных копий. Современная эволюционная биология смотрит неодобрительно на групповой отбор, отбор на уровне вида или сравнимой группы индивидов: естественный отбор происходит на более низком уровне, и все эволюционные тенденции, которые, как кажется, указывают на межвидовый отбор, обычно могут быть прослежены до последствий отбора на нижнем уровне. Фактически, если исключить особый случай родственного отбора, нет никаких отчетливых примеров адаптации, которая недвусмысленно шла бы на благо группе: в лозунге «на благо вида» нет никакого содержания. Проблема единицы отбора может быть выражена разными способами, так как отбор максимален на определенном уровне. На самом низком уровне бытия, на уровне атомов, не имеет значения, кто кого препарирует, ибо атомы переживают и убийцу, и нанесение увечий и резню. На много более высоком уровне, возьмем, к примеру, царство Животных, также не имеет значения, кто кого забивает, поскольку выживание царства не связано с его составом. Влияние на выживание становится гораздо более существенным, когда мы достигаем уровня индивидов и их генов, так как разница между убийцей и убитым теперь жизненно важна. Небольшой сдвиг к краю шкалы приводит нас к видам: смерть индивида определенно влияет на будущее вида, поскольку обычно лучше иметь столь много производителей, сколь это возможно, и ваше выживание является вкладом, при условии, что вы репродуктивно состоятельны. Класс млекопитающих также несколько более склонен к выживанию, когда обедающим является млекопитающее, а обедом — кто-то другой, но «собака съедает собаку» — и вообще, млекопитающее съедает млекопитающее факт, почти нейтральный. Сдвигаясь от индивидов к противоположному краю шкалы, мы сталкиваемся с их генами, которые одинаковы для индивида и для вида. Обладает ли обед чьими-то генами большей или меньшей важностью, чем обед просто кем-то? Одним из подходов к выявлению единицы отбора является установление сущности, которая является потенциально бессмертной. Бессмертными являются атомы, но они представляют царство минералов, а не царство организмов. Компоненты, из которых формируется двойная спираль ДНК («нуклеотидные основания», которые мы обсуждаем в главе 2), не несут жизни внутри, так же как буквы алфавита не являются литературой. Даже если бы эти компоненты были бессмертны, считать их живыми не приходится. Геном человека, полный набор ДНК в каждой клетке, также не бессмертен, поскольку он постоянно меняется в процессе, называемом мейотической рекомбинацией, который сопровождает половое воспроизведение, когда одна цепочка — генов — заменяется другой (обсуждение этой проблемы также содержится в главе 2). Тут мы меняем уровень, переходя к генам, репродуктивно активным цепочкам ДНК. Ген потенциально бессмертен — пока не подвергнется мутации — так как он переносится от генома к геному, от мыши к мыши, оставаясь практически неповрежденным.[3] Является ли тогда именно он единицей отбора? В своей книге Адаптация и естественный отбор (1966) Джордж Уильямс утверждал, что ген следует рассматривать как некоторую порцию хромосомного материала, потенциально сохраняющуюся в поколениях, число которых достаточно для того, чтобы считать ее единицей естественного отбора. Оксфордский зоолог Ричард Докинз (р. 1941) в своей заслуженно знаменитой книге Эгоистичный ген (1976) безжалостно развил эту идею и проследил, как посредством эгоистического действия ген разворачивается в биосистему и обеспечивает свое собственное выживание. Я упоминал в Прологе, что наука, как правило, углубляет свои прозрения и умножает свои успехи, переходя на более высокие уровни абстракции. Эта тенденция заметна и в биологии. Естественный отбор играет роль естественной кучи компоста для произрастания абстракций, а идентификация гена как единицы отбора явилась главным шагом в данном направлении. Докинз ищет поэтому естественный отбор, протекающий на самом низком уровне, в генах, и считает организм лишь временно используемым сосудом, который безжалостно эгоистичный (в техническом смысле, я подчеркиваю) ген арендует, чтобы обеспечить свое собственное распространение. Бессознательный ген формирует свой сосуд, его фенотип, так, чтобы наилучшим образом приспособить его к окружающей среде, ибо именно наиболее приспособленный сосуд будет гарантировать распространение гена. Но существует более низкий уровень отбора, еще более абстрактный, чем ген, потенциально даже более бессмертный. Ген кодирует фенотипическую информацию, такую как информация о строении тела, его окраске или физиологических модификациях, необходимых для усиления громкости рыка. Ген представляет собой физическую целостность, которая вынуждена обновляться, когда метаболические процессы копируют нити ДНК и обеспечивают поступление копий в каждую клетку и к следующему поколению. Сам по себе, как физическая целостность, даже ген не бессмертен, ибо физический ген должен непрерывно перестраиваться. Тот факт, что в ДНК закодирована информация, только деталь, ведь функция не есть основание. Но когда мы рассматриваем ген как единицу отбора, мы в действительности фокусируемся на информации, которую он передает, и точно так же как тело организма пригодно для использования в качестве сосуда для генов, так и последовательность элементов ДНК есть подходящая физическая реализация для информации, содержащейся в гене. Подлинно бессмертной компонентой жизни является не физический ген, ею является абстрактная информация, которую он содержит.[4] Информация бессмертна, и информация безжалостно эгоистична. Генетическая информация, вероятно, является предельной единицей отбора, с ДНК в качестве своей реализации и с телом в качестве сбрасываемого служебного сосуда. Живой мир возникает, когда неорганическое вещество спотыкается на пути, проходящем среди сложной, непредсказуемой информации, и обнаруживает, что может обеспечить бессмертие для этой информации, непрерывно воспроизводя ее. И здесь зарыта еще одна неистово бегущая Красная Королева, ибо постоянство достигается только вечным воспроизведением. Подобным же образом наш собственный номинально цивилизованный, культурный, разумный и рефлексирующий уровень жизни возник, когда организмы на пути, проходящем среди сложной, непредсказуемой информации, споткнулись о другие организмы, окружающие их и следующие за ними. Когда это случилось, был изобретен язык и все человеческие организмы, прошлые, настоящие и будущие, эффективно спаялись вместе в единый мегаорганизм с потенциально неограниченными возможностями. После этой риторической, но прочувствованной пышности слога самое время спуститься к проблемам пола. Одним из наиболее загадочных проявлений естественного отбора является эволюция полового воспроизводства. На первый взгляд пол выглядит хорошей идеей, в том смысле, что он наделяет виды генетической гибкостью и быстрой реакцией на меняющиеся условия. Однако здесь имеются проблемы. Во-первых, пол не является необходимым. Довольно многие виды превосходно обходятся без него. Партеногенез (непорочное зачатие) обычное дело у растений, где его более уместно называть партенокарпией. Мы уже упоминали партенокарпические одуванчики, но могли бы добавить и много других распространенных растений, таких как черная смородина (Rubus) и манжетка обыкновенная (Alchemilla). Неполовым путем размножаются и некоторые рептилии, наиболее заметны ящерицы Нового Света рода Cnemidophorus (семейство Teiidae), ящерицы Старого Света рода Lacerta (семейство Lacertidae) и слепозмейка (Raphotyphlops braminus; семейство Typhlopidae). Никакие млекопитающие неполовым путем не размножаются, если не считать противоположных утверждений из Библии. Во-вторых, пол неустойчив. Предположим, что некоторый вид размножается половым путем, и каждая пара производит многочисленное потомство, в котором половина мужских особей и половина женских. Чтобы численность популяции оставалась приблизительно постоянной, все потомство, за исключением примерно двух особей, должно погибнуть, чтобы остались в среднем одна мужская особь и одна женская. Предположим теперь, что с одной из женских особей случилась мутация, и она приобретает способность размножаться неполовым путем. Она снова произведет многочисленное потомство, из которого выживут примерно двое; однако они, являясь клонами матери, будут обе женского пола. Обе могут размножаться с помощью партеногенеза, производя еще больше самок. При условии, что одна асексуальная самка производит столько же потомков, сколько пара сексуальных партнеров (спорное предположение, разумеется, поскольку отцы часто выполняют определенные функции и после копуляции), через несколько поколений партеногенетическая женская популяция затопит исходную. Чтобы обеспечить устойчивость, у пола должно быть некое уравновешивающее преимущество. В-третьих, пол в высшей степени сложен. Половое воспроизведение зависит от загадочного механизма мейоза, в котором, как мы увидим в главе 2, число хромосом в зародышевых клетках (гаметах, сперме и яйцеклетке) делится пополам, но восстанавливается снова в соматических (типичных для тела) клетках после оплодотворения. Насколько же невероятно мощным должно быть селективное давление, чтобы привести к развитию столь «умного» механизма? Нет ничего необычного в развитии сложных механизмов путем сталкивания вместе и видоизменения уже существующих анатомических и биохимических свойств — примером являются бесчисленные независимые случаи эволюции глаза — но, как и само обладание глазом, пол был подарком, захватывающим дух, предложением, от которого организм не мог отказаться. Оксфордский биолог Уильям Гамильтон (1936-2000), которого Ричард Докинз считал кандидатом на титул самого выдающегося дарвиниста после Дарвина, полагал, что он идентифицировал этот подарок. Гамильтон глубоко интересовался паразитами, и незадолго до того, как он сам был иронически и трагически сражен одним из них, заболев малярией, предположил, что пол дает возможность организму оставаться на шаг впереди охотящихся на него паразитов. Совместная эволюция паразита и хозяина, в которой каждый из участников создает быстрое изменение среды для эволюции другого, нуждается в быстром и специфическом типе реакции, которую может обеспечить пол. Тщательный анализ динамики сосуществования, скорее похожего на маневрирование наций в период «холодной войны», показывает, что пол создает преимущество, обеспечивая механизм сохранения генетической информации, которая становится избыточной, но может быть востребована снова, когда генотип паразита возвращается к предшествующей инкарнации. Иными словами, пол создает хранилище для мечей в эпоху мушкетов, но мушкеты могут быть сняты с вооружения. Припрятанные мечи, однако, бесполезны, если мушкеты уступают место ядерному оружию; то есть пол бесполезен, если паразит скорее вырабатывает новую стратегию, чем возвращается к старой. Эта теория остается умозрительной, так как зависит от наличия специфической эволюционной связи между паразитом и хозяином, и ее трудно подтвердить экспериментально. Легче идентифицировать механизмы, которые поддерживают пол, чем механизмы, давшие начало этому сложному устройству. Во-первых, популяции с половым размножением более гибко приспосабливаются к окружающей среде, чем партеногенетические популяции. Так, благоприятные мутации могут произойти в обоих родителях по отдельности и даровать преимущество их потомкам, в партеногенезе одна мутация следует за другой. То есть в половых популяциях мутации могут происходить параллельно, а в неполовых только последовательно. Во-вторых, в половой популяции менее вероятно распространение вредных мутаций, потому что дефектные родители еще могут произвести нормального ребенка (это становится очевидным с точки зрения менделевской наследственности, глава 2), в то время как организмы, размножающиеся неполовым путем, могут избежать скверной мутации, только если тот же самый ген мутирует обратно, что невероятно. Половой диморфизм (различия во внешности самцов и самок одного и того же вида), в частности, экстравагантную внешность, которой часто отмечены самцы, тоже относительно легко объяснить или, по крайней мере, состряпать ему правдоподобное объяснение. Например, в королларии к своей теории эволюции пола Гамильтон считает, что чрезмерно пышный внешний вид самца является знаком того, что он здоров и свободен от паразитов. Исследование самца, приводимое самкой — то, что мы, люди, можем назвать «влюбленностью», — становится тогда похожим на медицинскую проверку. Пол, по-видимому, дарует преимущества на многих уровнях популяциям, индивидам и генам. Большая часть эволюционных сдвигов создает лишь малые преимущества: чтобы расплатиться за пол, преимущество должно быть огромным. И почему вообще должно возникнуть какое-то преимущество от смешивания генов кого-то вам незнакомого с вашими? Камнем преткновения здесь является то обстоятельство, что происхождение пола, например, продолжительность пути, который организмы должны пройти, чтобы до него добраться, остается тайной до сих пор. Давайте перейдем от ощущения, что Земля движется, к реально движущейся Земле. Никогда тектонические процессы не оказывали большего воздействия на наше собственное существование, чем при едва различимых переменах, которые произошли, когда африканская кора покрылась рябью, реагируя на давления, действовавшие на нее во время путешествия по южному полушарию. Около 20 миллионов лет назад поверхность Африки была по большей части плоской и покрытой на всем своем простирании тропическим лесом. Вы могли бы начать различать неоднородности лишь с момента 15 миллионов лет назад, когда локальное поднятие образовало плоскогорья из лавы, расположенные в областях, которые мы теперь называем Кенией и Эфиопией. Эти плоскогорья сформировались в чувствительной зоне, поскольку земля под ними начала разъезжаться. Когда щель между ними расширилась, плоскогорья обрушились, образовав глубокий и длинный разлом, Великую рифтовую долину, которая теперь простирается от современного Мозамбика через Эфиопию к Красному морю и далее, до самой Сирии. Вновь выросшие в результате этого возвышенности стали преградой для дождей в восточной части континента, и тропические леса постепенно выродились в открытую саванну. Теперь ландшафт создавал богатое разнообразие потенциальных сред обитания — несколько биомов — с влажными, жаркими, богатыми растительностью зонами в одних областях и сухими травянистыми пространствами в других. Для исследования оказались открытыми не только новые ниши, для исследования и освоения открылась и репродуктивная изоляция, ибо сообщества организмов были лишены возможности мигрировать через выросшие естественные барьеры. Организмы оказались в ловушке. Организмы оказались в ловушке физического, но не эволюционного пространства. Одним из важнейших следствий естественного отбора является существование Homo sapiens, вида, который сам H. sapiens считает апофеозом эволюции. Дарвин весьма чувствительно относился к выводам из своей теории, согласно которым человек скорее был потомком обезьян, чем отдельным творением. В стане христиан также были обеспокоены мыслью, что появление человека без участия грехопадения подрывает основание христианской церкви, центральной догмой которого является идея искупления первородного греха. Как бы то ни было, существуют недвусмысленные свидетельства того, что вы и я произошли от обезьяноподобных предков. Это происхождение столь важно для того, чтобы начать понимать самих себя и свое место в биосфере, что кажется уместным потратить некоторое время на его рассмотрение. В естественном отборе необходимо учитывать расселение (миграцию жизни на суше), двуногость (хождение вертикально на двух ногах, дающее рукам свободу манипуляций), энцефализацию (разрастание мозга относительно размеров тела) и возникновение культуры. И сегодня спорят, предшествовала ли двуногость расселению или следовала за ним. Одно из преимуществ двуногости в том, что она дает большую выносливость для преследования стад, и прямоходящее животное может дальше видеть через саванну и замечать хищников. Тем, кто думает, что принятие культуры — включая употребление орудий — послужило трамплином для нашего наступления, оппоненты доказывают, что первым шагом была энцефализация. Приматы обычно обитают на деревьях в тропических и субтропических лесных экосистемах. Мы распознаем их, отмечая характерные черты анатомии их рук и ног, способы их передвижения, возможности зрения, архитектуру их зубов и их интеллект. Эта последняя черта является центральной, поскольку первичной характеристикой приматов является эволюция интеллекта как образ жизни. Характеристики зубов важны, поскольку они позволяют нам установить тип диеты и, в частности, жило ли животное на деревьях, питаясь мягкими фруктами, или на земле, поедая более твердые семена и зерна. Приматы разбиты на две основные группы — полуобезьяны и антропоиды. Полуобезьяны включают лори и буш-беби; антропоиды — мелких обезьян, крупных обезьян и людей. 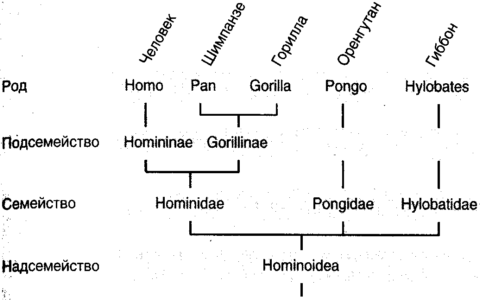
Рисунок 1.10 показывает генеалогическое дерево человека, и на нем стоит задержать взгляд не в последнюю очередь потому, что отдаленная человеческая история есть нечто, приводящее в замешательство и все еще требующее, чтобы его кусочки собрали вместе. Давным-давно, в эпоху Палеоцена («старую современную») Третичного периода (первоначально «третьей» эры) Кайнозойской («современной животной») эры ранние полуобезьяны разделились на современных полуобезьян, которые далее нас не касаются, и эволюционную ветвь, в будущем ведущую к нам. Во времена раннего Олигоцена (неподражаемого «мало современного») мелкие обезьяны Нового Света (платиррхины, что означает просто «с плоским носом», включающие в себя мартышек, ревунов и капуцинов) разбили лагерь в Южной Америке. Наша ветвь дерева отделилась позже, когда мелкие обезьяны Старого Света (катаррхины, что значит «с вдавленным носом», включающие живущих на деревьях толстотелов и живущих на земле макак и резусов) появились в Африке. Гоминиды впервые появились около 30 миллионов лет назад, в Миоцене («среднем современном») в форме дриопитеков (или «жителей дубов», что отражает их предполагаемое тропическое и субтропическое место обитания). Крупные обезьяны раннего Миоцена, такие как проконсул, по осанке были очень похожи на мелких обезьян: сам проконсул не имел хвоста (как крупные обезьяны), но держал тело параллельно земле (как мелкие обезьяны). Проконсул также имел противопоставленный большой палец, что является легким намеком на возможность использования им орудий. Ископаемых гоминидов находят почти во всех отложениях Миоцена в Африке и Евразии, и самыми ранними образцами является вид проконсул с возрастом в 22 миллиона лет. Примерно на половине пути через Миоцен, между 10 и 15 миллионами лет назад, дриопитеки, которых находят даже на севере вплоть до Испании и Венгрии, разветвились на несколько родов, которые включали Сивапитеков в Индии, Пакистане, а возможно, даже в Турции, и Рамапитеков в Африке. Сивапитеки, вероятно, предки орангутангов; Рамапитеков долго считали предками человека, но теперь думают, что они были просто одним из многих обезьяноподобных видов того времени. Гоминиды, жившие около 3 миллионов лет назад, разбились на две группы, одна с большим мозгом и маленькими коренными зубами, другая с маленьким мозгом и большими коренными зубами. Первая образовала род Homo; последняя — австралопитеков рода Austropithecus (южных обезьян). Первый образец последней группы, A. africanus, был обнаружен в 1924 г. при разработках карьера в Тунге, близ Йоханнесбурга, и казался предком Homo. Первоначально об этом была опубликована лишь небольшая заметка, главным образом потому, что свежие и болезненные воспоминания о пилтдаунской подделке породили скептицизм, и существовала точка зрения, что Африка неподобающее место для колыбели человечества; Англия гораздо предпочтительнее, лучше графства, прилегающие к Лондону, но юго-западные тоже сойдут. До сих пор не определено, в какое место филогенетического дерева следует поместить A. africanus. В 1962 г. Луис Лики, старейший из охотников за ископаемыми гоминидами, проводя раскопки в ущелье Олдуваи на равнине Серенгети в Танзании, наткнулся на останки гоминида, пользовавшегося орудиями, которого он определил как новый вид Homo habilis («человек умелый»), существовавший примерно 1,8 миллиона лет назад. Изготовление орудий из камня возникло, по-видимому, приблизительно в то же время, что и значительное развитие мозга, около 2,5 миллионов лет назад, и можно лишь предполагать, способствовало ли изготовление орудий росту мозга, или наоборот. Тогда H. habilis вызвал серьезные споры, и на том основании, что внутри вида бывает много физических вариаций, некоторые думали, что это большой A. africanus, в то время как другие полагали, что это маленький Homo erectus (с которым мы сейчас встретимся). Современная точка зрения, видимо, состоит в том, что H. habilis является самостоятельным видом или, по крайней мере промежуточным видом на пути к H. erectus. Современная точка зрения, кажется, состоит также в том, что несколько скелетов, соединенных вместе в виде H. habilis, представляют на самом деле два отдельных вида, H. habilus и Homo rudolfensis, последний (названный в честь озера Рудольф, где он был найден, теперь озеро Туркана в северной Кении) имел немного более крупный мозг и более современную структуру мозга. До сих пор неизвестно, какой из этих видов в конце концов привел к H. sapiens, поскольку «очевидный» ответ, что это H. rudolfensis, приходит в противоречие с наблюдениями, согласно которым он имеет не те видимые анатомические черты, которые для этого необходимы. Теперь (около 2 миллионов лет назад) и здесь (в Африке) на сцену выступает Homo ergaster («человек работающий»), обремененный комплектом своих каменных инструментов. Он выше своих предшественников и имеет более крупный мозг. Почти полный скелет «турканского мальчика» (рис. 1.11), найденный Ричардом Лики в 1984 г. на западном берегу озера Туркана является представительным: его сохранность позволяет предположить, что он, вероятно, утонул примерно 1,6 миллиона лет назад, поскольку не был разорван на куски плотоядными животными. Homo ergaster осваивал широкие открытые пространства и умеренный сухой климат; он пользовался орудиями, которые классифицируются как Ашельские. Это название происходит от Сент-Ашель во Франции, где была найдена коллекция относительно усовершенствованных двусторонних каменных орудий, таких как каплеобразный ручной топор. Предполагают, что H. ergaster был скорее охотником, чем собирателем. 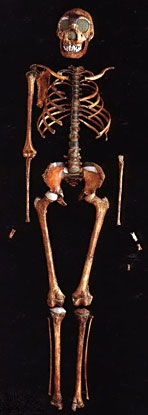
Около 1,8 миллиона лет назад H. ergaster распространился в Азию, и здесь, как и в Африке, эволюционировал в H. erectus («человек прямоходящий»), который уже полностью ходил вертикально на двух ногах и был, возможно, так же высок, как современные люди, но с более тяжелым лицом, низким лбом и надбровными дугами (рис. 1.12). Предполагается, что H. erectus следовал за стадами, которые, в свою очередь, следовали за отступлением лесов, вызванным ледниковым охлаждением планеты, и сопровождал их через тропические и субтропические зоны Саудовской Аравии до юга Центрального Китая. Отсюда он перебрался по перешейку на Яву, где в 1891 г. Эжен Дюбуа в ходе своих поисков «пропущенного звена» нашел на реке Соло части «яванского человека». 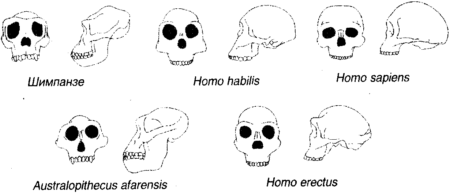
В соответствии с «африканской гипотезой» H. sapiens («человек разумный»), по-видимому, возник в Африке 150 тысяч лет назад, возможно, от H. ergaster, но, возможно, и от не столь яркого (с точки зрения скудости свидетельств о размерах его мозга) H. erectus. В свою очередь, H. sapiens пришлось конкурировать с H. neanderthalensis, образцом «пещерного человека», чьи останки нашли рабочие каменоломен в 1856 г. в Фельдерхофер Гротто над рекой Дюссель в долине Неандер в Германии (рис. 1.13). 
Неандертальцы имели более крупные тела, были более мускулистыми, более коренастыми и с более бочкообразной грудной клеткой, чем мы, и хотя их мозг был в среднем крупнее, чем мозг современного человека, в их черепах, как полагают некоторые комментаторы, ощущался недостаток фронтальных долей. Они проявили способность адаптироваться к холодному климату. Позднее их образцы находили на Ближнем Востоке, в Азии и Северной Африке. Их технология обработки камня сегодня классифицируется как Мустьерская, по имени местоположения пещер в Ле Мустье во Франции. В дальнейшем, возможно, в результате контактов с H. sapiens около 40 тысяч лет назад они заимствовали идеи из Ашельской технологии и усовершенствовали ее до версии, известной как Шательперронская, по имени соответствующего района пещер. Орнаменты, сделанные из оленьих рогов и костей, позволяют сделать предположение о том, что между этими двумя видами имела место торговля. Проблеском, осветившим домашний быт неандертальцев, является находка флейты, датируемой временем от 43 до 67 тысяч лет назад, и, возможно, неандертальского происхождения, с отверстиями, соответствующими диатонической шкале из семи нот в современной человеческой музыке. Помимо музыкальных вечеров неандертальцы имели развитую социальную организацию, сотрудничали на охоте, возможно, общались с помощью языка (хотя строение их гортани указывает на то, что их язык страдал недостатком артикуляции) и хоронили своих покойников, что можно интерпретировать как чувствительность (но можно и как обрушение свода). С другой стороны, они могли быть каннибалами — что можно было бы также объяснить чувствительностью, буквальным принятием в себя тех, кого любишь, — если отбросить наше культивируемое отвращение. Если отдельный предмет искусства, одинокий отполированный и продырявленный зуб мамонтенка, является той отправной точкой, из которой могут быть сделаны надежные антропологические экстраполяции, то неандертальцы были артистичны. Артистичностью они обладали, а вот жизнестойкостью нет, ибо H. neanderthalensis исчезли около 30 тысяч лет назад, став тупиком эволюции, сухим листом на ветке дерева жизни. Вымирание неандертальцев началось около 40 тысяч лет назад с волны вторжения, которая хлынула через Европу с востока на запад и закончилась около 27 тысяч лет назад. Завоевателями были кроманьонцы (рис. 1.14), разновидность H. sapiens, почти идентичная современному человеку и названная по имени места Кро Маньон в центральной Франции (близ деревни Лес Эйзи в Дордони), где в 1868 г. были найдены первые образцы. Кроманьонец был адаптирован к теплу, и это подтверждает точку зрения, что он (вместе со своей «ею», разумеется) прибыл из Африки. Кроманьонцы сметали все перед собой, возможно благодаря своему значительному превосходству в орудиях (т.е. в оружии), известных как Ориньякские, которые включали кость и олений рог, а также изысканную глиняную посуду, или благодаря тому, что у неандертальцев не было защиты от принесенных ими болезней, или потому, что из-за недостаточно артикулированной речи неандертальцы не могли достаточно эффективно общаться, чтобы координировать свою оборону. Кроманьонцы жили в шалашах и употребляли ловушки для животных, луки и стрелы, черенки и рукоятки для ножей и битум для их крепления. Война теперь вступила на путь индустриализации, и этому суждено было стать проклятьем человеческой истории на все последующие тысячелетия. 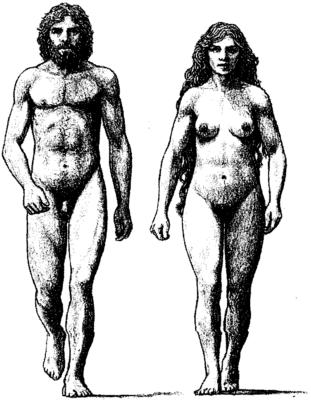
Современные люди, однако, когда не заняты бойней, имеют теперь все необходимое, чтобы поразмышлять о своей окружающей среде, о своей физической и психологической природе, о строении вещества, которое их окружает и которое они медленно познают, чтобы подчинить своей воле. Что будет дальше, зависит от того, куда приведут эти начинания. Примечания:1 Построенный из дерева гофер, ковчег был 300 локтей в длину, 50 локтей в ширину и 30 локтей в высоту; локоть — длина руки от локтевого сгиба до кончиков пальцев, около 45 сантиметров). 2 «Бигль» имел 27 метров в длину и 7 метров в ширину в средней части судна, что примерно эквивалентно 15×4 «дарвинов». 3 Я говорю «практически неповрежденным», потому что даже если во время мейоза, происходящего в середине гена, возникнет случайная поломка ДНК, шаг рекомбинации восстановит этот ген в новом геноме. 4 Не совсем так, энтропия в конце концов уничтожает и информацию. — Прим. пер. |
|
||
| Главная | В избранное | Наш E-MAIL | Добавить материал | Нашёл ошибку | Наверх | ||||
|
|
||||